Genetische Kontrolle der Rumpfentwicklung in der Maus
Der Rumpf und der Schwanz entwickeln sich durch kontinuierliche Verlängerung der vorderen Körperanlage, die Kopf und Hals hervorbringt. Dieser Verlängerungsprozess wird durch eine kaudale Struktur gesteuert, den Primitivstreifen gefolgt von der Schwanzknospe. Diese fungieren als Wachstumszone, in der Stamm- und Vorläuferzellen ständig Nachkommen produzieren, die die verschiedenen Gewebe generieren.
Drei Hauptzelllinien werden während der Rumpfbildung fortlaufend aus Vorläuferzellen gebildet, aus denen fast alle Gewebe und Organe entstehen. Dies sind das Mesoderm, das Neuroektoderm und das Endoderm. Die Mesodermderivate erzeugen die meisten Gewebe des Rumpfes, sie bilden z.B. das Skelett, die Skelettmuskulatur, die Gliedmaßen, die Körperwand, die Blutgefäße, die Nieren, die Gonaden und die glatten Muskeln. Das Neuroektoderm bildet das Rückenmark und das Endoderm den Mittel- und Enddarm.

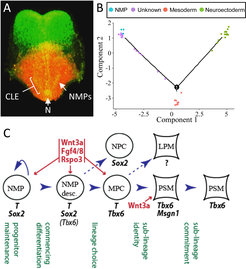
Das Neuroektoderm und das Mesoderm stammen von neuromesodermalen Vorläuferzellen (NMPs) ab, einem Zelltyp mit begrenzter Fähigkeit zur Selbsterneuerung. NMPs werden durch Sox2 und Brachyury kontrolliert, Transkriptionsfaktoren, die für die neurale und mesodermale Entwicklung wesentlich sind. Die Selbsterneuerung von NMPs hängt von einer selbstregulierenden Rückkopplungsschleife zwischen dem sezernierten Induktormolekül Wnt3a und Brachyury ab. Dieses Genpaar bildet das Schlüsselkontrollmodul für die Rumpf- und Schwanzentwicklung. Zusammen kontrollieren sie die Erhaltung der Vorläuferzellen, die axiale Verlängerung zur Bildung von Rumpf und Schwanz, die Mesodermbildung und das anfängliche dorso-ventrale Muster der erzeugten Gewebe.
Die NMP-Zellteilung erzeugt Nachkommen, die sich differenzieren können, um das Gewebe des Rumpfes zu bilden. Diese frühen Nachkommen müssen sich entscheiden, welches Schicksal sie wählen, entweder Richtung Rückenmark oder Richtung mesodermale Gewebe. Diese Wahl wird durch antagonistische Aktivitäten zwischen Sox2 und Brachyury bestimmt, bei denen jeder Faktor die Kontrollfaktoren der eigenen Stammlinie hochreguliert und die Kontrollfaktoren der konkurrierenden Stammlinie unterdrückt. Die große Mehrheit der NMP-Nachkommen wird von WNT-Induktion zusammen mit Brachyury dominiert und wählt daher ein Schicksal Richtung Mesodermentwicklung. Mesodermale Zellen werden weiteren Stammentscheidungen in Richtung auf paraxiales, intermediäres oder laterales Schicksal unterzogen. Die Wahl des paraxialen Schicksals, die zur Bildung des axialen Skelett- und Skelettmuskelapparats führt, wird durch die WNT-Signalisierung zusammen mit dessen Zieltranskriptionsfaktoren Brachyury, Tbx6 und Msgn1 gesteuert.
Die Bildung lateraler und ventraler Mesodermgewebe ist weniger gut verstanden und wird zurzeit in unserem Labor untersucht. Der zelluläre Kern und die Kontrollmechanismen, die an der Schwanzentwicklung beteiligt sind, sind ebenfalls noch unklar. Handelt es sich nur um eine Verlängerung des Rumpfes, an der dieselben NMPs und Notochord-Vorläufer beteiligt sind? Und was ist der zelluläre Ursprung des Notochords in Rumpf und Schwanz? Dies sind zwei der dringlichsten Fragen, die zurzeit in unserem Labor untersucht werden.